微信扫一扫,访问移动端
×
首页
关于我们
公司简介
发展历程
平台展示
组织架构
资质荣誉
知识产权
企业文化
医学科研服务
测序分析服务
细胞生物学实验
分子生物学实验
动物学实验
组织病理学实验
其他平台
临床前CRO服务
药物筛选研究
药物有效性评价
动物疾病模型构建
药物安全性评价
药代动力学研究
资源中心
经典案例
合作伙伴
行业新闻
报告下载
技术文献
联系我们
联系方式
在线留言
招贤纳士
资源中心
您所在位置:
首页
>>
资源中心
>>
行业新闻
经典案例
合作伙伴
行业新闻
报告下载
技术文献
通过eIF2α激酶信号通路调节饥饿和病毒诱导的自噬
时间:2021-09-14 01:43:00
点击量:937次
摘要
eIF2α激酶是一类进化上保守的丝氨酸/苏氨酸激酶,可调节应激诱导的翻译停滞。在这里,我们表明,酵母eIF2α的激酶,GCN2,Gcn2p的目标磷酸化位点,eIF2α的的丝氨酸51和eIF2α的调节转录激活因子,GCN4,是另一个基本的应激反应,饥饿诱导自噬是必不可少的。哺乳动物IFN诱导的eIF2α激酶,PKR,在GCN2破坏的酵母和pkr中拯救饥饿诱导的自噬null和Ser-51非磷酸化突变eIF2α鼠胚胎成纤维细胞在由单纯疱疹病毒感染引发的自噬中存在缺陷。此外,PKR和eIF2αSer-51依赖性自噬被单纯疱疹病毒神经毒力蛋白ICP34.5拮抗。因此,自噬是病毒毒力基因产物靶向的eIF2α激酶途径的一种新的进化保守功能。
研究背景
在营养匮乏和病毒感染期间,细胞需要一种策略来合成必需蛋白质,以应对由环境耗竭或细胞内寄生引起的有限供应。该策略不仅必须涉及严格的翻译调控机制,还必须涉及从现有蛋白质生成新的氨基酸库。因此,在营养限制条件下,真核细胞通过自噬途径同时降低整体蛋白质合成并增加蛋白质降解率,这一过程涉及自噬溶酶体对细胞内容物的大量降解。尚不清楚应激诱导的翻译抑制和应激诱导的自噬是否受共同或遗传不同途径的调节。
真核起始因子2-α(eIF2α的)在Ser-51通过eIF2α的蛋白激酶的保守家族的磷酸化是在应激诱导的翻译调控,但的应激诱导的调节机制自噬还不是很清楚。在真核细胞的氨基酸饥饿期间,自噬是生存所必需的,并且已经鉴定出几种对自噬至关重要的酵母和哺乳动物APG和AUT基因。至少在酵母中,这些基因在雷帕霉素(TOR)信号通路的自噬抑制靶标下游起作用。然而,对于启动自噬过程必不可少的上游细胞基因知之甚少。此外,尚不清楚氨基酸饥饿以外的刺激,如病毒感染、内质网应激或血红素耗竭,这些触发应激诱导的翻译停滞是否也会触发应激诱导的自噬。
本研究提出了假设,即eIF2α激酶,即应激诱导的翻译控制程序的充分表征的调节剂,也参与了应激诱导的自噬的调节,并且激活eIF2α激酶依赖性翻译停滞的特定应激刺激也激活了eIF2α激酶依赖性自噬。为了评估eIF2α激酶在自噬调节中的作用,本文研究了在eIF2α激酶信号通路中具有基因工程突变的氮饥饿酵母细胞和病毒感染和氨基酸饥饿哺乳动物细胞中的自噬。
方法与结果
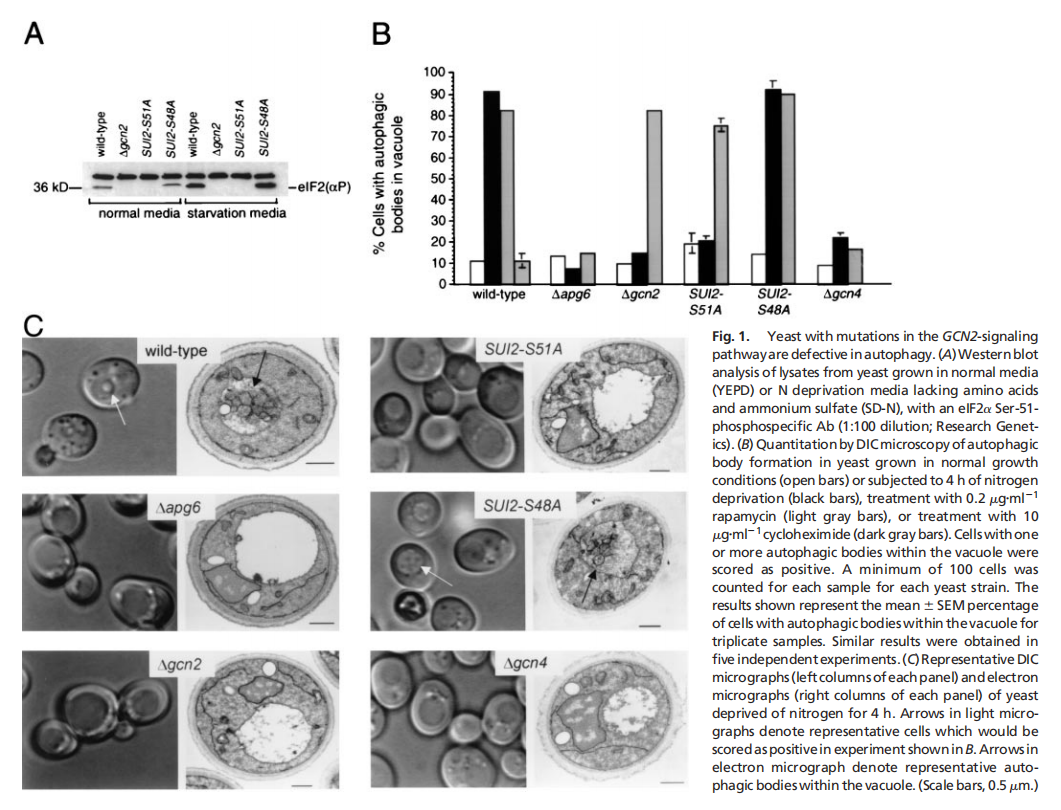
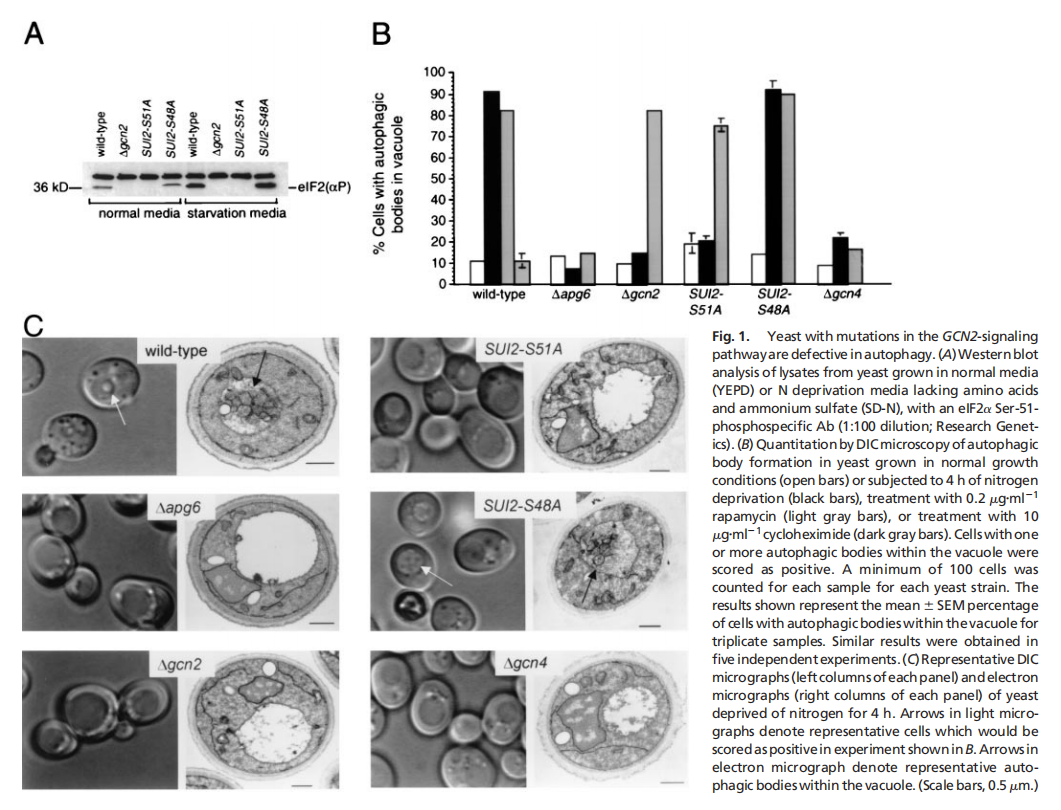
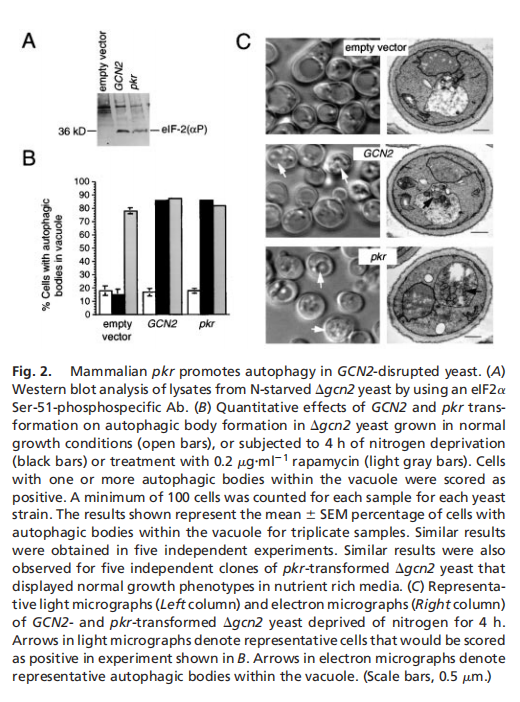
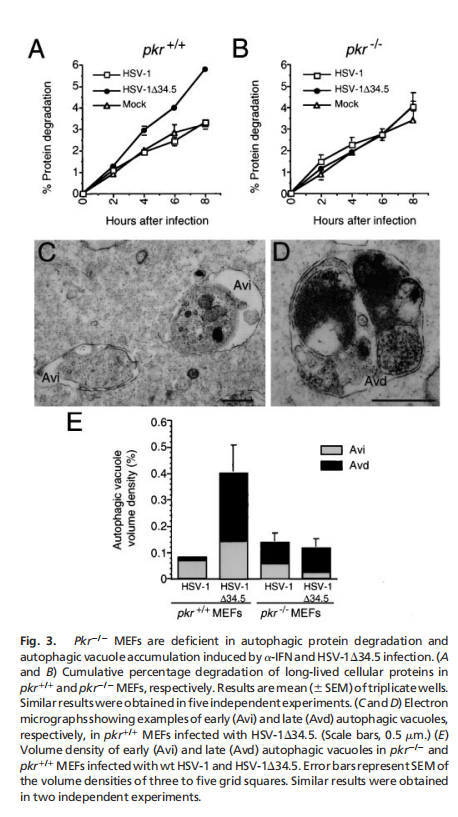
实验结果表明,酵母eIF2α激酶信号通路对于饥饿诱导的自噬是必不可少的,eIF2α激酶、GCN2及其磷酸化靶位点eIF2α的Ser-51,对于酵母中饥饿诱导的自噬至关重要。GCN2依赖的转录反激活因子GCN4,而不是转录抑制,对酵母中饥饿诱导的自噬至关重要。酵母在突变GCN2 -7号信令通路是细胞自噬缺陷(见图一);哺乳动物IFN诱导的eIF2激酶PKR促进gcn2破坏的酵母菌的自噬(见图二);PKR是病毒诱导的mef自噬所必需的,它被HSV-1编码的PKR抑制剂和神经毒性蛋白ICP34.5阻断(见图三);eIF2Ser-51是mef中病毒诱导和氨基酸饥饿诱导的自噬所必需的(见图四)。
这些数据表明,哺乳动物细胞中病毒和氨基酸饥饿诱导的自噬都需要eIF2α激酶底物位点eIF2αSer-51。需要进一步研究以确定哺乳动物GCN2,如酵母GCN2,是否是在饥饿诱导的自噬过程中作用于eIF2α上游的eIF2α激酶。目前尚不清楚,其他刺激,如内质网应激和血红素缺乏,分别激活eIF2α激酶、PERK和HRI(21),是否在哺乳动物细胞中诱导eIF2αSer-51依赖性自噬。
结论
总之,我们的数据表明酵母和哺乳动物eIF2α激酶途径在自噬调节中具有新颖的、进化上保守的作用。我们已经表明,酵母和人类eIF2α的激酶,GCN2和PKR,酵母和人类磷酸化位点由eIF2α的激酶靶向,eIF2α的丝氨酸51,和酵母GCN4转录反式激活因子,对于应激诱导的自噬都是必不可少的。此外,我们的数据表明,已知刺激eIF2α激酶依赖性翻译停滞的不同压力刺激(例如,营养剥夺和病毒感染)也会刺激eIF2α激酶依赖性自噬。众所周知,自噬是一种重要的细胞防御机制,可以抵御这些压力刺激之一,即营养缺乏。我们的观察表明自噬受PKR调节,PKR是一种特征很好的抗病毒分子,并且自噬被HSV-1神经毒力基因产物拮抗,这表明自噬也是一种重要的细胞防御机制,可以抵御病毒感染。在营养缺乏和病毒感染期间,eIF2α激酶信号通路对翻译和自噬的同步调节可能是使真核细胞成功适应环境压力的基本机制。作为其毒力策略的一部分,病毒可能会颠覆这种宿主适应性机制。